Structure and gating in plant potassium channels
Structure and gating in plant potassium channels
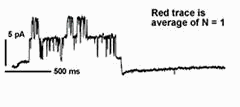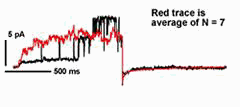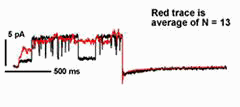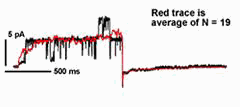 In many cases, transport of K+ across the plant plasma membrane is dominated by two K+ channels. Of these, current through the outward-rectifying K+ channels shows an unusual voltage sensitivity: gating of the channel interacts with [K+] outside so that the net K+ flux through the channel is always directed outward. This dependence on extracellular K+ for gating makes good "physiological sense" in an environment where the concentration of the cation often varies by more than 100-fold. It also raises fundamental biological questions about the mechanism of K+ sensing. Kinetic analysis has shown that 2 K+ ions interact with the channel to gate its activity in Vicia guard cells, and at a site (or sites) remarkably similar in its physico-chemical properties to the channel pore [Blatt and Gradmann (1997) J. Membr. Biol. 158,241].
In many cases, transport of K+ across the plant plasma membrane is dominated by two K+ channels. Of these, current through the outward-rectifying K+ channels shows an unusual voltage sensitivity: gating of the channel interacts with [K+] outside so that the net K+ flux through the channel is always directed outward. This dependence on extracellular K+ for gating makes good "physiological sense" in an environment where the concentration of the cation often varies by more than 100-fold. It also raises fundamental biological questions about the mechanism of K+ sensing. Kinetic analysis has shown that 2 K+ ions interact with the channel to gate its activity in Vicia guard cells, and at a site (or sites) remarkably similar in its physico-chemical properties to the channel pore [Blatt and Gradmann (1997) J. Membr. Biol. 158,241].
This K+-sensitivity is found in the yeast YKC1/TOK1 K+ channel and in the Shaker-like SKOR and GORK K+ channels of Arabidopsis. It is intrinsic to the channel proteins, since the same characteristics are observed when the channels are heterologously expressed. It is of interest that the structures of these channels is very different. Unlike SKOR and GORK, the YKC1 K+ channel is unique in having a tandem pair of pore domains and an N-terminal structure of four transmembrane-helices similar to the Shaker family of voltage-gated K+ channels. Indeed, it appears that these different K+ channels have evolved very different molecular mechanisms that underpin their common physiological behaviour. We previously identified two putative binding sites for K+ near the extracellular mouth of the channel [Vergani, et al. (1998) EMBO J. 17,7190; Vergani and Blatt (1999) FEBS Lett. 458,285] with distinct kinetic effects on the gating process. The situation of these sites in a cleft on the outer surface of the channel, and the effects of mutations around these sites from work ongoing [see Johansson and Blatt (2006) Biochem J. 393,645], strongly suggests that K+ binding in the cleft stabilises closed conformations of the channel.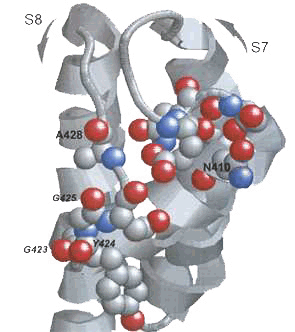
Our work with the SKOR and GORK K+ channels - part of a collaboration with Drs. I. Dreyer and B. Müller-Röber (University of Potsdam) - has yielded a very different picture of the mechanics of K+ sensitivity in the plant K+ channel and provides an intriguing example of parallel evolution. In this case, K+ -dependent gating is associated with intramembrane sites and it appears to arise from interactions analogous to the 'foot-in-the-door' model of Clay Armstrong [see Yellen (2002) Nature 419,35]. However, gating in the plant K+ channels flies in the face of conventional thinking: it seems that K+ occupation of the pore actually promotes closure of the gate [Johansson, et al. (2006) Plant J, in press]! We are now preparing to explore this mechanism as an essential step towards a general understanding of K+channel gating.