SNARE function and redundancy in plant membrane traffic
SNARE function and redundancy in plant membrane traffic

In plants, directed vesicle traffic is essential for maintaining cell polarity, growth and development, and is responsible for delivery of some of the most biologically interesting and commercially important products, including various alkaloids, anticancer drugs, dyes and enzymes [cf. Walker (2003) Plant Physiol. 132,44]. These are highly dynamic processes involving the rapid turnover of large areas of membrane surface. In some cell types, notably pollen, vesicle traffic may drive the turnover of plasma membrane surface area in excess of 0.01 cm2 per minute at the growing tip. For such seamless integration of membrane traffic, the cell must match vesicles with their destinations to ensure efficient fusion and delivery of specific membrane proteins and soluble cargo. These functions are carried out by the superfamily of membrane and membrane-associated proteins known as SNAREs (soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptors). Subsets of SNAREs occur at vesicle and target membranes and interact to form a tetrameric bundle of coiled helices which draws the membrane surfaces together and thus provides a mechanism for recognition, docking and fusion [see Pratelli et al. (2004) Trends Plant Sci. 9,187]. Functional SNARE complexes comprise one element of each of four sub-motif domains, designated Qa, Qb, Qc and R with reference to the core amino acid of the SNARE submotif [see Bock, et al. (2001) Nature 409,839]. This combinatorial model for SNARE interaction goes some way to explaining their high specificity, but also raises questions about their redundancy, especially in plants.
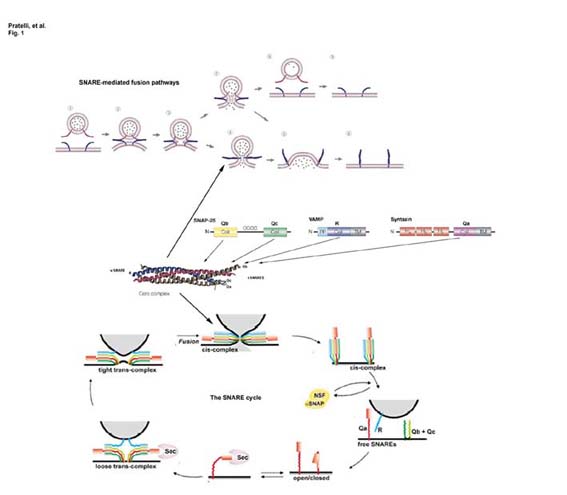
Do plant SNAREs function in vesicle trafficking? Almost certainly yes. However, to date there is still little direct evidence to support specific functions of these proteins in vivo. Our work has demonstrated that the cytosolic domain of the tobacco SNARE NtSyp121 acts to suppress traffic of a secreted Green Fluorescent Protein (secGFP) marker in vivo [Geelen, et al. (2002) Plant Cell 14,387]. Expression of the NtSyp121 cytosolic domain leads to an accumulation of secGFP in the ER and Golgi, and a cessation of growth which is rescued by overexpression of the full-length NtSyp121 protein. The same cytosolic domain (and that of AtSyp121) affects traffic of the Arabidopsis KAT1 K+ channel but, intriguingly, not of the PMA2 H+-ATPase to the plasma membrane [Sutter, et al. (2006) Plant Cell, in press]. These observations are most easily understood if binding of the cytosolic domain forms non-functional SNARE complexes and thereby suppresses vesicle traffic to the plasma membrane. They also demonstrate that plasma membrane traffic post-Golgi passes via at least two different pathways.
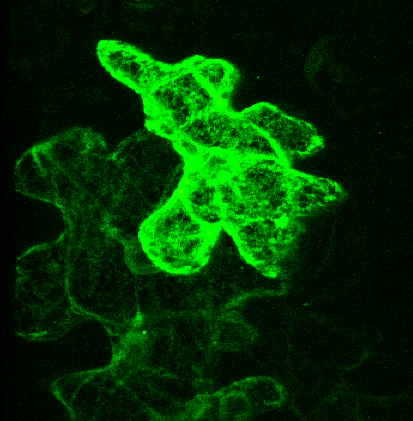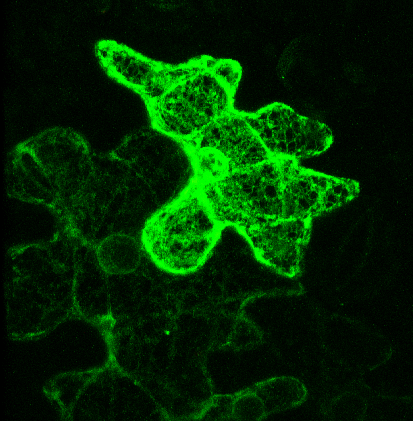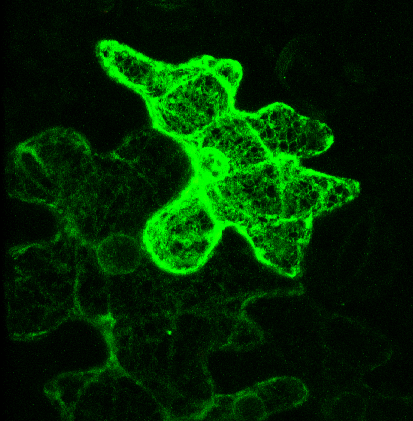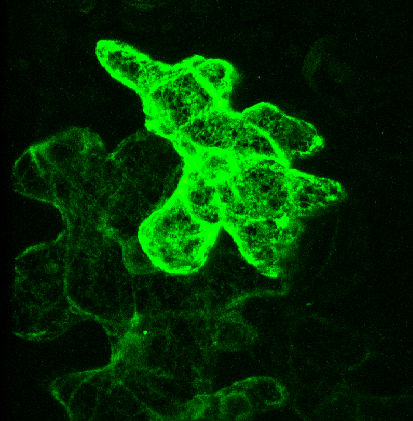 Pictured right - secGFP accumulated in the ER with expression of a soluble (dominant-negative) fragment of NtSyp121 [see Geelen, et al. (2002)]
Pictured right - secGFP accumulated in the ER with expression of a soluble (dominant-negative) fragment of NtSyp121 [see Geelen, et al. (2002)]
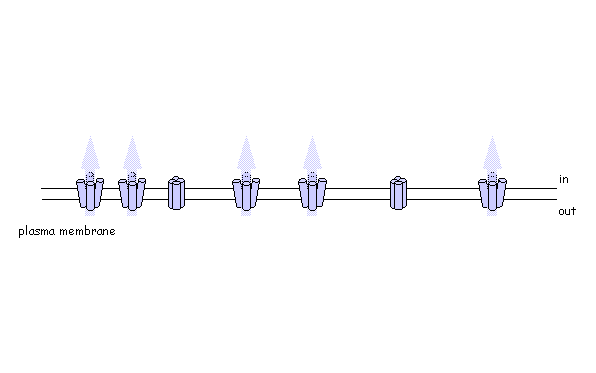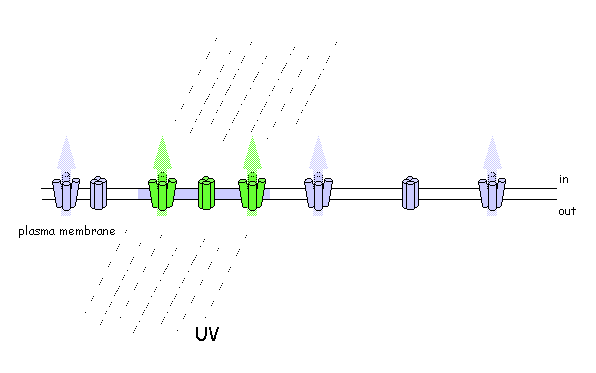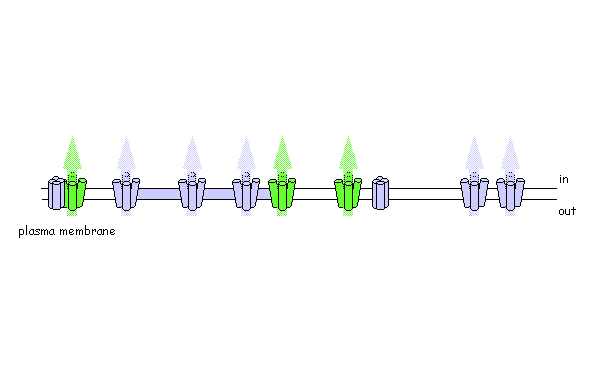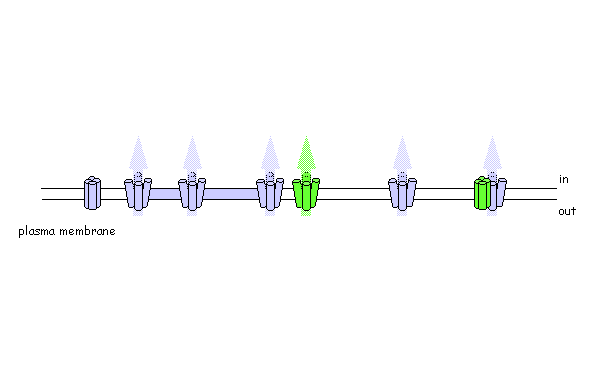
Are plant SNAREs functionally specialised? The genome sequence of Arabidopsis shows a remarkable diversity of SNARE proteins (68 in Arabidopsis compared with 35 in humans, 21 in yeast and 20 in Drosophila) as well as several unique gene subfamilies. It is likely that new functions underlie this diversity of proteins, but assigning biological roles is difficult, even where close homologies do occur across phylogenetic boundaries. We are exploring SNARE function, taking advantage of advanced tagging technologies, including use of photo-activatable GFP, to monitor the traffic of specific soluble and intrinsic membrane proteins. To test SNARE function, we are currently manipulating traffic using a combination of dominant-negative and rescue strategies in vivo [Geelen, et al. (2002)] that faithfully recapitulate the known interactions (and lack thereof) between several plasma membrane SNAREs. Among other findings, these studies have recently uncovered some surprising differences in the patterns of traffic among membrane ion transport proteins, including K+ channels [Sutter, et al. (2006) Plant Cell, in press].
Using photo-activatable GFP to monitor lateral membrane protein movement