

Roles for SNAREs in signalling
Roles for SNAREs in signalling
Roles for [Ca2+]i - and possibly also pHi - may also extend to coupling membrane vesicle traffic at the plasma membrane with regulated ion channel activities. Because stomatal movements (and plant cell expansion generally) entail very large changes in volume and surface area, it is essential that the guard cell maintains tight control over membrane flow to and from the plasma membrane. The mechanics of vesicle traffic in this case almost certainly depends on SNARE proteins for vesicle docking and fusion, and these proteins are likely also to contribute to other aspects of membrane function and control.
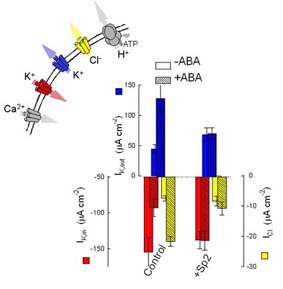 Two new developments have now placed membrane trafficking squarely in the limelight of stomatal physiology, ABA signalling, and highlight the importance of coordinating ion transport with membrane traffic in ion channel control. First, recent studies of exocytosis and endocytosis in Vicia guard cell protoplasts have confirmed an active turnover of membrane during osmotic challenge [Homann (1998) Planta 206,309; Kubitscheck, et al. (2000) Planta 210,423] and its coupling to the number of active channels in the membrane [Meckel, et al. (2004) Plant J. 39,182]. These results point to a highly dynamic process of membrane cycling that is responsive to membrane tension. The second development has come from our identification of the tobacco and Arabidopsis syntaxins (Q-SNAREs) NtSyr1 and AtSyr1 (NtSyp121 and AtSyp121 [Leyman, et al. (1999) Science 283,537; Leyman, et al. (2000) Plant J. 24,389], the first plasma membrane-localised SNAREs to be described in plants. The NtSyr1 gene was pulled out of an expression cloning screen for an ABA receptor. We found that the protein is required for ABA-mediated control of both the K+ and Cl- channels of the guard cell plasma membrane, as a soluble (dominant-negative) peptide - the so-called Sp2 fragment - blocked ABA action in regulating all three ion channels.
Two new developments have now placed membrane trafficking squarely in the limelight of stomatal physiology, ABA signalling, and highlight the importance of coordinating ion transport with membrane traffic in ion channel control. First, recent studies of exocytosis and endocytosis in Vicia guard cell protoplasts have confirmed an active turnover of membrane during osmotic challenge [Homann (1998) Planta 206,309; Kubitscheck, et al. (2000) Planta 210,423] and its coupling to the number of active channels in the membrane [Meckel, et al. (2004) Plant J. 39,182]. These results point to a highly dynamic process of membrane cycling that is responsive to membrane tension. The second development has come from our identification of the tobacco and Arabidopsis syntaxins (Q-SNAREs) NtSyr1 and AtSyr1 (NtSyp121 and AtSyp121 [Leyman, et al. (1999) Science 283,537; Leyman, et al. (2000) Plant J. 24,389], the first plasma membrane-localised SNAREs to be described in plants. The NtSyr1 gene was pulled out of an expression cloning screen for an ABA receptor. We found that the protein is required for ABA-mediated control of both the K+ and Cl- channels of the guard cell plasma membrane, as a soluble (dominant-negative) peptide - the so-called Sp2 fragment - blocked ABA action in regulating all three ion channels.
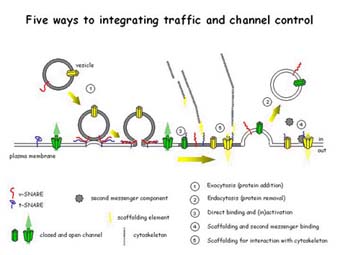 Our subsequent work has confirmed that NtSyr1 is essential for secretion, and contributes to cellular growth and development throughout the plant as well as for stomatal response to ABA and drought stress [Geelen, et al. (2002) Plant Cell 14,387]. Thus, NtSyr1 appears to play dual roles in vesicle trafficking and in ABA signal transduction. One hypothesis is that NtSyr1 functions either as a scaffolding protein, or a second messenger/modulator to set the poise of one or more signalling elements in the membrane. Alternatively, our results may point to a critical role for vesicle trafficking in the (necessarily rapid!) transfer of selected plasma membrane ion channels between pools located on the plasma membrane and on intracellular compartments. Our recent findings [Sutter, et al. (2006) Plant Cell, in press; see also Sutter, et al. (2006) Traffic, in press] have shown that NtSyr1 and AtSyr1 are important for anchoring K+ channels within the plane of the plasma membrane. These observations support the idea of a role in scaffolding and raise questions about the importance of position in the membrane for channel regulation (LOCATION! LOCATION! LOCATION!). Other important questions now are whether ABA and hydrostatic pressure changes recruit common sets of vesicles or influence the lateral distributions of ion channels, whether different vesicle pools contain different proteins, and whether SNAREs such as NtSyr1 and AtSyr1 find roles in seeding, formation and endocytotic traffic of vesicles from the plasma membrane as well as in anchoring K+ channels in the membrane.
Our subsequent work has confirmed that NtSyr1 is essential for secretion, and contributes to cellular growth and development throughout the plant as well as for stomatal response to ABA and drought stress [Geelen, et al. (2002) Plant Cell 14,387]. Thus, NtSyr1 appears to play dual roles in vesicle trafficking and in ABA signal transduction. One hypothesis is that NtSyr1 functions either as a scaffolding protein, or a second messenger/modulator to set the poise of one or more signalling elements in the membrane. Alternatively, our results may point to a critical role for vesicle trafficking in the (necessarily rapid!) transfer of selected plasma membrane ion channels between pools located on the plasma membrane and on intracellular compartments. Our recent findings [Sutter, et al. (2006) Plant Cell, in press; see also Sutter, et al. (2006) Traffic, in press] have shown that NtSyr1 and AtSyr1 are important for anchoring K+ channels within the plane of the plasma membrane. These observations support the idea of a role in scaffolding and raise questions about the importance of position in the membrane for channel regulation (LOCATION! LOCATION! LOCATION!). Other important questions now are whether ABA and hydrostatic pressure changes recruit common sets of vesicles or influence the lateral distributions of ion channels, whether different vesicle pools contain different proteins, and whether SNAREs such as NtSyr1 and AtSyr1 find roles in seeding, formation and endocytotic traffic of vesicles from the plasma membrane as well as in anchoring K+ channels in the membrane.