Calcium signalling and ion channel regulation in guard cells
Calcium signalling and ion channel regulation in guard cells
Ca2+ signalling and ion channel regulation in guard cells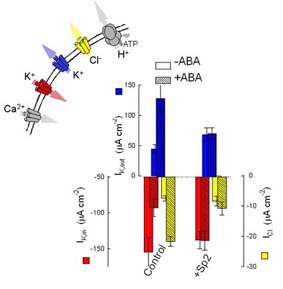
Guard cells open and close stomata by taking up and releasing solutes (mainly KCl and other K+ salts) which drives water movement across the plasma membrane. Both K+ and Cl- channels participate in these events and their activities are coordinately regulated. Under conditions of water stress, for example, abscisic acid (ABA) activates the Cl- channels and one class of so-called outward-rectifying K+ channels to promote net KCl loss; it also suppresses the activity of a second class of (inward-rectifying) K+ channel that normally mediates K+ uptake. Many aspects of how ABA, and other environmental stimuli, coordinate these changes in membrane transport have been well-characterised, but others remain a mystery, and remarkable new discoveries continue to surface.
At least two parallel signalling pathways operate in the guard cells, one mediated through cytosolic-free [Ca2+] ([Ca2+i) and the other through cytosolic pH (pHi). Protein phosphorylation also contributes to ion channel control. Both the [Ca2+]i and pHi pathways contribute to K+ and Cl- channel control during stomatal closure. A rise (and, more recently, evidence of oscillations) in [Ca2+]i has long been associated with control of the inward-rectifying K+ channel. Our work [Grabov and Blatt (1998) PNAS 95,4778] demonstrated that [Ca2+]i transients in guard cells are coupled to transitions in plasma membrane voltage. Membrane hyperpolarisations trigger a wave of high [Ca2+]i that passes centripetally through the cell. These [Ca2+]i transients are triggered by Ca2+ entry across the plasma membrane and led to our identification of inward-rectifying Ca2+ channels and their modulation by ABA [Hamilton, et al. (2000) PNAS 97,4967] and protein phosphorylation [Köhler and Blatt (2002) Plant J.32,185]. All indications are that ABA targets a site closely associated with the Ca2+ channels at the plasma membrane. Surprisingly, our attempts to identify an ABA receptor turned up something completely different (see Roles for SNAREs in signalling).

An important question is how do [Ca2+]i and membrane voltage integrate K+ and Cl- channel control. Our working hypothesis [Hamilton, et al. (2000); Blatt (2000) Ann. Rev. Cell Dev. Biol. 16,221] proposes a role for [Ca2+]i and voltage as a signalling 'cassette' that drives the membrane through oscillations of solute uptake and loss. The time-averaged balance between uptake and loss determines the net flux and steady-state solute content within the cell. Stomatal closure (and, conversely, opening) is achieved by altering the threshold, or set-point, for transitions between these two activities. Indeed, evidence of defined ranges of [Ca2+]i oscillations encoding stomatal movements [Allen, et al. (2001) Nature 411,1053] fits neatly within this concept. The model proposes experimentally testable predictions, some of which have already been borne out. We have confirmed (1) that the [Ca2+]i rise in ABA requires membrane hyperpolarisation and that ABA ‘tunes’ the interaction between [Ca2+]i and voltage [Grabov and Blatt (1998) PNAS 95,4778], and (2) that the inward-rectifying K+ channels are exquisitely sensitive to [Ca2+]i within a narrow concentration range just above normal resting values [Grabov and Blatt (1999) Plant Physiol. 119,277]. Propagation of the [Ca2+]i signal internally is also controlled by ABA. In this case, the effect arises from a synthesis of nitric oxide which, in turn, promotes intracellular Ca2+ release via a cADPR-dependent pathway [Garcia-Mata, et al. (2003) PNAS 100,11116].
Continuous recording of [Ca2+]i during two voltage ramps (-50 to -200 mV) before and after treatment with the phosphatase antagonist calyculin A. After Köhler and Blatt (2002).
Work from this laboratory was also the first to identify a role for pHi in ABA-mediated control of K+ channels. In ABA, guard cell pHi rises by as much as 0.3 to 0.5 units over periods of 5-10 min to promote the activity of K+ channels for K+ efflux [Blatt and Armstrong (1993) Planta 191,330]. Under some conditions, pHi can rise by more than 0.5 units in 20-30 seconds [Thiel, et al. (1993) PNAS 90,11493] – a rise at least as dramatic as that of [Ca2+]i ! However, pHi and [Ca2+]i function independently in vivo [Grabov and Blatt (1997) Planta 201,84]. The pHi signal is important for K+ channel regulation mediated by the ABI1 protein phosphatase [Armstrong, et al. (1995) PNAS 92,9520; Leube, et al. (1998) FEBS Lett 424,100]. How these changes in pH come about is still one of the big puzzles of guard cell biology.
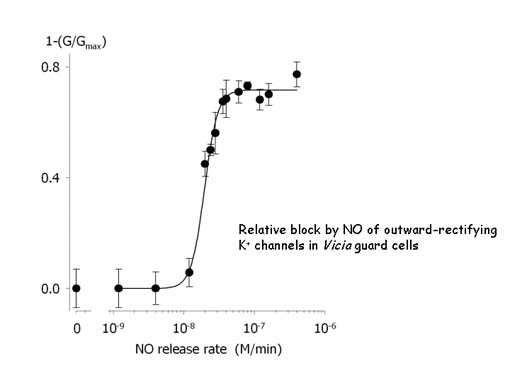
Our recent explorations into the action of nitric oxide (NO) have turned up yet another level of complexity to guard cell K+ channels, parallelling recent interest in their regulation by reactive oxygen species. At moderate elevation (20-50 nM), NO suppresses current through the same K+ channels mediating K+ efflux. All evidence suggests the effect involves S-nitrosylaton of cysteine residues closely associated with the K+ channels. The physiological significance is less clear at present, but the observations may be related to the effects of oxidative stress in suppressing stomatal closure [see Willmer and Fricker (1996) Stomata (Longman, New York)] and, in some circumstances in promoting stomatal opening [Black and Black (1979) J. Exp. Bot 30,291].